Authors Bob Thoolen Henk Koster Anton van Kolfschoten Martin de Haan
Abstract
Descriptions of two rare gastric neuroendocrine tumors (carcinoids) of enterochromaffin (ECL) cells in CD-1 mice (2/50) from a 104-week oncogenicity study of a serotonergic/dopaminergic compound are presented. These tumors were detected at necropsy and confirmed by histopathology in hematoxylin and eosin- and Chromogranin A-stained slides. ECL cell counts of the glandular stomachs were determined by quantitative image analysis and did not reveal any hyperplastic changes as possible predisposing lesions for carcinoid formation. To investigate the possibility of drug-induced hypergastrinemia as the cause of tumor formation of ECL cells, gastrin blood levels were measured after treating mice for 7 days with the test substance. In this study, Omeprazole, the positive control, raised gastrin levels, while the test material did not. It was concluded that these two tumors were an example of “late-life”-occurring, spontaneous neuroendocrine tumors in the stomachs of aged CD-1 mice.
Keywords
Carcinoids; CD-1 mice; Chromogranin A; ECL cells; gastrin; neuroendocrine tumor; stomach.
Introduction
In a 2-year oncogenicity study with a serotonergic/dopaminergic compound in CD-1 mice, gross examination showed in the glandular stomachs of two male mice raised areas that appeared to be neuroendocrine tumors at microscopy. The question was whether these tumors were drug-induced or spontaneously occurring tumors. From a battery of test results (Ames, gene mutation, chromosome aberration, DNA repair, and micronucleus tests), it was concluded that the compound was nongenotoxic. Additional techniques and a mechanistic study were performed to clarify the situation.
Neuroendocrine tumors (carcinoids) are rare in both humans and animals (8, 13, 18, 28, 32), accounting for less than 1% of all gastrointestinal tumors in man. In humans, they are also seen in association with the multiple endocrine neoplasia (MEN) syndromes, most commonly with type I. Since APUD cells (cells active in the amine precursor uptake and decarboxylation), from which these tumors arise, are widespread, carcinoid tumors are found in a variety of locations, mostly in the appendix or terminal ileum but sometimes in the pancreas, gallbladder, ovary, or testis. As in animals, carcinoids of enterochromaffin cells (ECL cells) occur in humans after long-term hypergastrinemia in conditions like pernicious anemia and gastrinoma (14, 33). ECL cells of the stomach are APUD cells and are the predominant endocrine population of the glandular stomach. ECL cells produce histamine and respond to gastrin by the release of this secretory product. ECL cells play a crucial role in the physiology of gastric secretion, and hyperplasia and tumors of these cells are a well-documented consequence of long-term hypergastrinemia (1, 11, 33, 35).
Little information is available on spontaneous carcinoids in rodents. Drug-induced hyperplasia of enterochromaffin cells and neuroendocrine tumors are reported for both rats and mice after treatment with ICI 162846, Omeprazole, Cimetidine, Ranitidine, Loxtidine, and Ciprofibrate (2– 4, 6, 7, 9, 12, 27, 30, 34). This article describes two neuroendocrine tumors (carcinoids) in a mouse oncogenicity study that was part of the preclinical safety program. The occurrence and pathogenesis of these tumors are discussed in light of additional investigations on the ECL cells and the available scientific and historical background data.
Materials and Methods
Mouse Carcinogenicity Study
Three groups (groups 3–5) of 50 male and 50 female CD-1 mice received the test substance via the diet for 104 weeks at dose levels of 3.8, 7.5, and 15.0 mg/kg/day for males and 7.5, 15.0, and 30.0 mg/kg/day for females. Two control groups (groups 1 and 2) of 50 animals/sex of the same strain received the untreated basal diet. In addition, three groups that received the same dosage levels as above were maintained for toxicokinetic assessment. At termination, a full necropsy was performed on all animals, including histopathological evaluation. The in-life experimental procedures, which were undertaken during the course of all studies, were subjected to the provision of the United Kingdom Animals (Scientific Procedures) Act of 1986.
Additional Investigations
To investigate the possibility of drug-induced preneoplastic hyperplasia of ECL cells, morphometric analysis was performed on the stomachs of all mice. Tissues were embedded in paraffin wax, and sections were cut at 4 µm and stained with hematoxylin and eosin. The stomachs were examined using Chromogranin A stain and image analysis for ECL-cell hyperplasia (21, 25, 38). Chromogranin was demonstrated by subjecting sections of stomach to an antibody against chromogranin, followed by a linking antibody, and then a streptavidin complex conjugated with peroxidase. This peroxidase was visualized by the diaminobenzidine reaction. The Chromogranin A-positive cells (CAS cells) were counted using a Seescan image analysis system. The parameter measured was the number of Chromogranin-positive cells per the length of mucosa.
Statistical Analysis of CAS-Positive Cells
Statistical analysis was performed for the number of Chromogranin A-positive cells per length of mucosa. Means and standard deviations were calculated. The data were tested for their underlying distribution using Shapiro– Wilk normality testing of the residuals after fitting a two-factor (sex and group) preliminary analysis of variance (ANOVA) model. It was found necessary to apply a square-root transformation to the data, which is common for cell count data. One-way analysis of variance, with group as the factor, was then applied to the transformed data for each sex separately. If the group effect was significant at the 5% level, ordinary t-tests were performed to compare each group with the others. Significance testing was carried out at the 5% and 1% levels only. The covering ANOVA avoided the need to apply multiple comparison adjustments to the t-tests.


7-Day Dietary Study of Mouse Gastrin Secretion
To assess the effect of the test substance on gastrin levels in blood, mice were treated by continuous dietary administration for 7 days. An untreated group and a reference (positive control) group dosed with Omeprazole (proton pump inhibitor) were included. Omeprazole is known to induce high levels of gastrin in the blood by blocking the production of stomach acid, and as such was used to demonstrate an increase in gastrin levels (23). At day 7, blood samples were taken from all groups twice. Mice were housed two per cage, 12/sex/treatment group. The dose levels chosen were the same as those of the carcinogenicity study, with an extra dose group of 48 mg/kg/day for both sexes. Positive control groups of both sexes received Omeprazole by oral gavage at 138 mg/kg/day. For mice receiving the test substance in the diet, subgroup A was sampled at 10.00 a.m. and subgroup B was sampled at midnight. For mice receiving Omeprazole by oral gavage, subgroup A was sampled 2 h after dosing and subgroup B was sampled 12 h after dosing. On day 7, blood samples were collected into heparinized tubes, containing aprotinin as a stabilizing agent, for the measurement of plasma gastrin levels (pg/ml) by radioimmunoassay using a Diagnostic Product test kit (Los Angeles, CA). The statistical analyses of the data were analysis of variance followed by Student’s t-test for the positive control group and Williams’s test for dosed animals to test a dose response (23).
Results
During the overall treatment (week 0 to 104), both male and female body weight gains were reduced by 16% in the top dose groups when compared to controls. In the female mice there was minimal adipose tissue observed at this dose at gross pathology. At histopathology, there was a dose-related increase in uterine atrophy, mammary hyperplasia and neoplasia, and pituitary tumors. A no-effect level was not determined in females. No treatment-related nonneoplastic changes were observed among male mice. For the neoplastic changes, two neuroendocrine tumors were noted in the glandular region of the stomach in two males receiving 15 mg/kg/day.
Histopathology Neuroendocrine Tumors
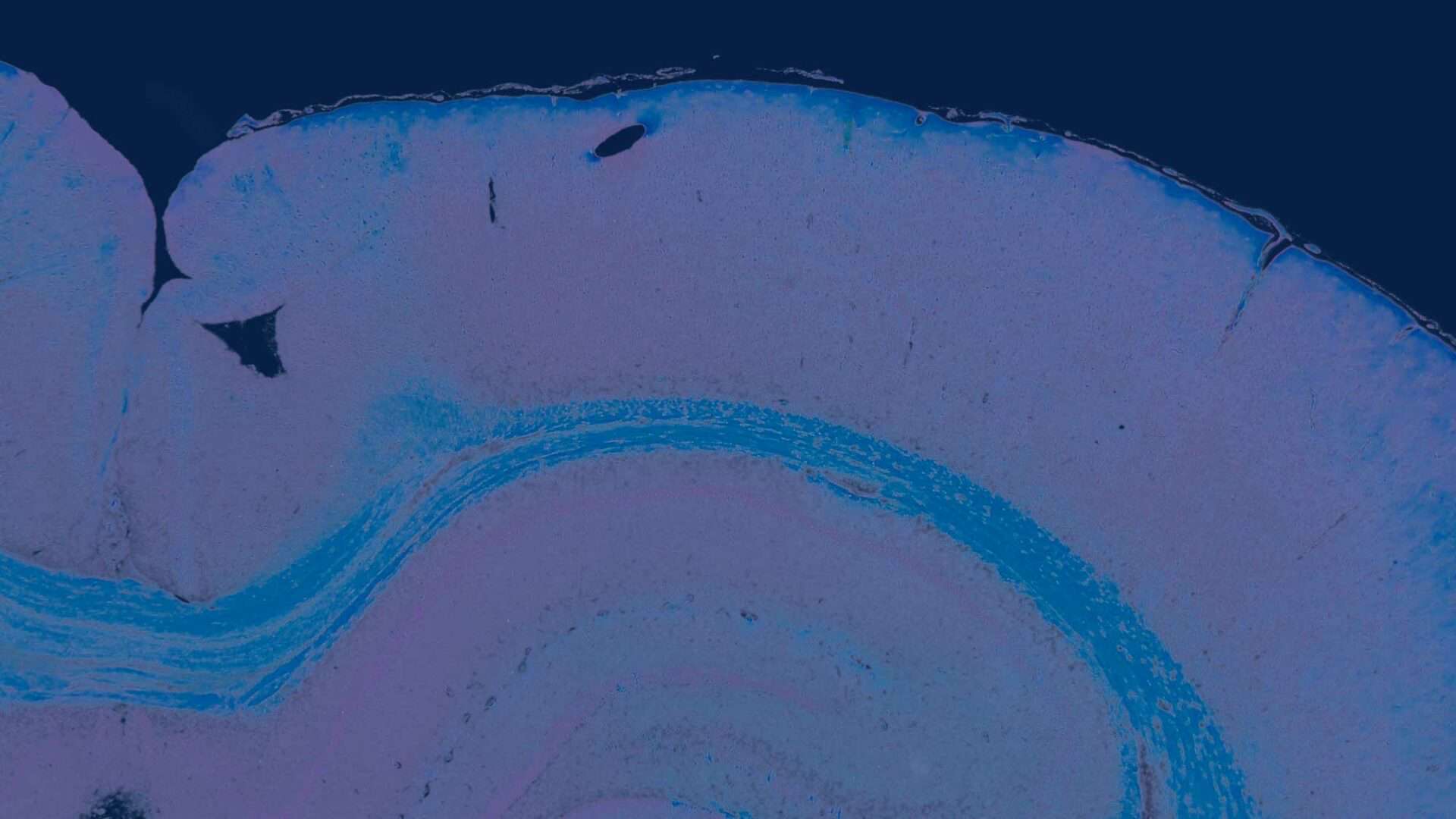
In each of two male mice there was a gastric neuroendocrine tumor confirmed by additional immunohistochemical staining with Chromogranin A (Figures 1 and 2).
Case 1 (Figure 1A) was from a male at terminal kill. The tumor had a solid growth pattern localized in the glandular mucosa and extended below the muscularis mucosa. Occasional small “oat-cell” nests of neuroendocrine cells were also evident beneath the muscularis mucosa. Tumor cells showed slight to moderate pleomorphism with enlarged euchromatic nuclei, scarce mitotic figures, and immunoreactivity for chromogranin (Figure 1, B and C).
Case 2 (Figure 2A) was from a male at terminal kill. A tumor invaded through the muscularis mucosa as a solid nodular growth largely located in the submucosa, with a few scattered nests of tumor cells at the periphery (Figure 2A). Solid cords of tumor cells were present in the mucosa (Figure 2A). As in case 1, tumor cells show slight to moderate pleomorphism with enlarged euchromatic nuclei, scarce mitotic figures, and immunoreactivity for chromogranin (Figure 2B).
The results of the image analysis of Chromogranin A-positive stained cells (CAS) in sections of the stomach mucosa after statistical analysis (20, 29, 36, 37) are presented in Figure 3. There was no dose-related change in gastric ECL cell density. For males, group 3 had statistically lower means than groups 1, 2, 4, and 5 (p < .01, p < .01, p < .05, and p < .01, respectively). Group 4 had a lower


mean than group 2 (p < .01), but there was no change at the highest dose (Figure 3). For females there was no statistical difference between groups.
Effect of Test Substance on Blood Gastrin
The analysis of gastrin blood levels revealed decreased gastrin levels among treated males (Figure 4) sampled at midnight and in treated females (Figure 5) sampled at 10:00 a.m. However, there was no clear dose-related trend, except perhaps for males at midnight, and there was high individual variation. Data indicated that the test substance when dosed at up to 48 mg/kg/day resulted in a slight but not consistent reduction in gastrin levels in the blood and did not certainly cause hypergastrinemia in mice. In addition, the decreased groups mean values did not follow a clear dose-related trend, and there was a degree of individual variation apparent in the gastrin levels that was reflected in the differences noted between the groups. The intergroup variation and the difference between sexes were considered to be due mainly to the disparity in feeding patterns of mice at the time points measured, rather than due to treatment. Among sexes treated with Omeprazole, sampled at 10:00 a.m. (time point A), the animals had values that were considered to be both comparable with those of the control. However, the positive control group (Omeprazole, 138 mg/kg/day) revealed increased gastrin secretion among both sexes sampled at midnight (time point B), with the female mean values attaining statistical significance.
DISCUSSION
In a 2-year oncogenicity study, neuroendocrine tumors were identified in two male CD-1 mice. ECL cell counts were

performed on stomachs to detect any treatment effects on predisposing hyperplastic changes. There were no treatment effects on the number of ECL cells. However, because of the rare occurrence of these kinds of tumors in mice (18, 19, 22), historical tumor data from both Charles River U.K. (vendor) and United States as well as National Toxicology Program (NTP) tumor data were evaluated. Neuroendocrine tumors are rare in rodents. They are most commonly caused by antisecretory drugs (3, 7, 12, 27, 31, 34) and are more common in rats than mice. This higher incidence in rats is probably because rats are more endowed with ECL cells, the cell type constituting the tumors (9). The mechanism of induction of carcinoids has been referred to as the “gastrin hypothesis” (3, 5, 7, 10– 12, 17, 35). The proposed sequence of events is gastric acid blockade hypergastrinemia ECL cell hyperplasia carcinoid. ECL cell hyperplasia and neoplasia are consequences of long-term continuous hypergastrinemia. The induction of these tumors in rodents via this mechanism does not pose a risk to humans, as available evidence indicates that widely used potent antisecretory drugs, such as Omeprazole and Loxtidine, are not carcinogenic for human. The lack of hyperplasia of stomach ECL cells casts doubt on a critical role for hypergastrinemia in this case, since hyperplasia has been observed in all other studies where neuroendocrine tumors occurred.
In the 7-day study, only the Omeprazole group showed marginal/slight hypergastrinemia, which was not seen in the animals treated with the test substance at dosages comparable to the carcinogenicity study. Therefore, the occurrence of the two neuroendocrine tumors could not be correlated with hypergastrinemia. In addition, neuroendocrine tumors rarely occur in control mice in standard 1.5-year CD-1 mouse oncogenicity studies. However, the chance of detecting this type of tumor is greater in extended 2-year studies (3, 9). Background data on the spontaneous incidence of gastric neuroendocrine tumors in 2-year studies are scarce, and a single case, with metastasis to regional lymph nodes, was found in B6C3F1 mice (18, 22). With regard to the occurrence of neuroendocrine tumors in rodents, Praomys (Mastomys) natalensis is known as the rodent with a high genetic susceptibility for the development of carcinoid tumors due to hypergastrinemia and is used as an experimental model in this respect (15, 16, 24). Few spontaneous gastric carcinoid tumors were found in striped field mice (Apodemus agrarius) (26), and eight cases were reported in NTP studies (19).
We conclude that there was no relationship to treatment because of the following reasons: Such tumors were not seen in male or female BioF1D Alexander Syrian hamsters, which were also treated with the test substance for 89 weeks (females) and 101 weeks (males). No carcinoids or other neuroendocrine tumors were found in other organs. In addition, there were no preneoplastic changes. The tumors occurred only in one sex and one species (oncogenicity study was also done in hamsters); there were no relevant preneoplastic changes and no hypergastrinemia. These tumors should not be considered treatment-related. Rather they are rare spontaneous occurrences. This article presents the first description of spontaneous occurrence of gastric carcinoid tumors in CD-1 mice.
Acknowledgements
We are grateful to Dr. C. Gopinath, Dr. P. Chambers, Dr. P. Ryle, Dr. Robert Harling, and all technical staff from Huntingdon Life Sciences for their collaboration and conduct of the studies. We also thank Andre Giesken and Marijke Mulder for technical assistance in preparing the manuscript. Lastly, we thank Bert Pagel and Rene de Bruijn, Weesp, who were involved in creating the photomicrographs.
References
- Belaiche J, Delwaide J, Louis E, Gast P (1993). Enterochromaffin-like cells and their receptors: Physiological role and physiopathological significance. Acta Gastroenterol Belg 56: 3– 4.
- Betton GR, Dormer CS, Wells T, Pert P, Price CA, Buckley P (1987). Fundic mucosal ECL hyperplasia and carcinoids in rodents following chronic administration of the histamine H2-receptor antagonist SK&F 93479 and other antisecretory agents. Toxicol Pathol 15: 365.
- Betton GR, Dormer CS, Wells T, Pert P, Price CA, Buckley P (1988). Gastric ECL-cell hyperplasia and carcinoids in rodents following chronic administration of H2-antagonists SK&F 93479 and oxmetidine and omeprazole. Toxicol Pathol 16: 288–98. 10.1177/019262338801600222
- Colin-Jones DG, Langman MJS, Lawson DH, Vessey MP (1985). Post-marketing surveillance of the safety of cimetidine: Mortality during second, third and fourth years of follow-up. Br Med J 291: 1084–1088. 10.1136/bmj.291.6502.1084
- Delle Fave G, Helander H, Holt S, Modlin IM, Powers R, Solcia E, Soll A, Tielemans Y, Wright NA (1994). Acid suppression and gastric mucosal cell biology. Dig Dis Sci 39: 1843–52. 10.1007/BF02088113
- Eissele R, Arnold R (1993). Risiko der Entwicklung von Tumoren unter der Therapie mit dem Protonenpumpen-Inhibitor Omeprazol (Risk for developing tumors in therapy with the proton pump inhibitor Omeprazole). Versicherungsmedizi n 45(4): 126–129.
- Ekman L, Hansson E, Havu N, Carlsson E, Lundberg C (1985). Toxicological studies on omeprazole. Scand J Gastroenterol Suppl 108: 53–69. https://doi.org/10.3109/00365528509095819
- Emminger A, Mohr U (1982). Tumours of the oral cavity, cheek pouch, salivary glands, oesophagus, stomach and intestines. In: Pathology of Tumours in Laboratory Animals—Tumours of the Hamster, Turusov V (ed). IARC scientific publications, Lyon, pp 45–50.
- Greaves P (1990). VII. Digestive system 1. In: Histopathology of Preclinical Toxicity Studies—Interpretation and Relevance in Drug Safety Evaluation, Elsevier, Amsterdam, pp 321–325.
- Hakanson R, Chen D, Andersson K, Monstein HJ, Zhao CM, Ryberg B, Sundler F, Mattsson H (1994a). The biology and physiology of the ECL cell. Yale J Biol Med 67: 3– 4. PMC2588926
- Hakanson R, Chen D, Tielemans Y, Andersson K, Ryberg B, Sundler F, Mattsson H (1994b). ECL cells: Biology and pathobiology. Digestion 55 (suppl 3): 38–45. 10.1159/000201200
- Havu N, Mattsson H, Ekman L, Carlsson E (1990). Enterochromaffin-like cell carcinoids in the rat gastric mucosa following long-term administration of Ranitidine. Digestion 45: 189–195. 10.1159/000200245
- Head KW (1990). Tumours of the alimentary tract. In: Tumors in Domestic Animals, Moulton JE (ed). University of California Press, Berkeley, pp 45–50.
- Kokkola A, Sjoblom SM, Haapiainen R, Sipponen P, Puolakkainen P, Jarvinen H (1998). The risk of gastric carcinoma and carcinoid tumours in patients with pernicious anaemia—A prospective follow-up study. Scand J Gastroenterol 33: 88–92. 10.1080/00365529850166266
- Kolby L, Wangberg B, Ahlman H, Modlin IM, Granerus G, Theodorsson E, Nilsson O (1996). Histidine decarboxylase expression and histamine metabolism in gastric oxyntic mucosa during hypergastrinemia and carcinoid tumor formation. Endocrinology 137: 4435–4442. 10.1210/endo.137.10.8828505
- Kolby L, Wangberg B, Ahlman H, Modlin IM, Nilsson O (1998). Histamine metabolism of gastric carcinoids in Mastomys natalensis. Yale J Biol Med 71: 207–215. PMC2578997
- Larsson H, Hakanson R, Mattsson H, Ryberg B, Sundler F, Carlsson E (1988). Omeprazole: Its influence on gastric acid secretion, gastrin and ECL cells. Toxicol Pathol 16: 267–72. 10.1177/019262338801600220
- Leininger JR, Jokinen MP (1994). Tumours of the oral cavity, pharynx, oesophagus and stomach. In: Pathology of Tumours in Laboratory Animals— Tumours of the Mouse, Turusov V, Mohr U (eds). IARC Scientific Publications, Lyon, pp 167–194.
- Leininger JR, Jokinen MP, Dangler CA, Whiteley LO (1999). Oral cavity, esophagus, and stomach. In: Pathology of the Mouse, Maronpot RR, Boorman GA, Gaul BW (eds). Cache River Press, Vienna, IL, pp 42–45.
- Levene H (1960). Robust tests for equality of variances. In: Contributions to Probability and Statistics, Olkin et al. (eds). Stanford University Press, Stanford, CA, pp 278–292.
- LIoyd RV (1999). Immunohistochemical markers of endocrine/neuroendocrine tumors. J Histotechnol 22(3): 231–237. https://doi.org/10.1179/his.1999.22.3.231
- Maekawa A, Enomoto M, Hirouchi Y, Yamakawa S (1996). Changes in the upper digestive tract and stomach. In: Pathobiology of the Aging Mouse, Mohr U, Dungworth DL, Capen CC, Carlton WW, Sundberg JP, Ward JM (eds). ILSI Press, Washington, DC, pp 267–286.
- Mattsson H, Andersson K, Hakanson R, Larssson H, Sundler F, Wallmark B (1988). Sex and species differences in plasma gastrin and gastrin-dependent variables. Studies in rats and mice during Omeprazole treatment. Gastroenterology 94(5): 2.
- Nilsson O, Wangberg B, Johansson L, Modlin IM, Ahlman H (1992). Praomys (Mastomys) natalensis: A model for gastric carcinoid formation. Yale J Biol Med 65: 741–751. PMC2589773
- Oberg K, Janson ET, Eriksson B (1999). Tumour markers in neuroendocrine tumours. Ital J Gastroenterol Hepatol 31(2): 160–162.
- Oh SW, Chae C, Jang D (1997). Spontaneous gastric carcinoids tumors in the striped field mouse (apodemus agrarius). J Vet Med Sci 59: 703–706. 10.1292/jvms.59.703
- Poynter D, Selway SAM, Papworth SA, Riches SR (1986). Changes in the gastric mucosa of the mouse associated with long-lasting unsurmountable histamine H2 blockade. Gut 27: 1338–1346. PMC1434077 or 10.1136/gut.27.11.1338
- Rubin E, Farber JL (1994). The gastrointestinal tract. In: Pathology. J. B. Lippincott, Philadelphia, pp 668–670.
- SAS Institute, Inc. (1990). SAS/STAT User’s Guide. Version 6, 4th ed. SAS Institute, Inc., Cary, NC.
- Spencer AJ, Barbolt TA, Henry DC, Eason CT, Sauerschell RJ, Bonner FW (1989). Gastric morphological changes including carcinoid tumors in animals treated with a potent hypolipidemic agent, ciprofibrate. Toxicol Pathol 17(1): 7–15. 10.1177/01926233890171P102
- Streett CP, Robertson JL, Crissman JW (1988). Morphologic stomach findings in rats and mice treated with the H2 receptor antagonists, ICI 125,211 and ICI 162,846. Toxicol Pathol 16: 299–304. 10.1177/019262338801600223
- Takahashi M, Hasegawa R (1990). Tumours of the stomach. In: Pathology of Tumours in Laboratory Animals—Tumours of the Rat, Turusov V, Mohr U (eds). IARC Scientific Publications, Lyon, pp 129– 135.
- Waldum HL, Brenna E, Sandvik AK (1998). Relationship of ECL cells and gastric neoplasia. Yale J Biol Med 71: 325–335. PMC2578987
- Waldum HL, Kvetnoi IM, Sylte R, Schulze B, Martinsen TC, Sandvik AK (1998). The effect of the peroxisome proliferator ciprofibrate on the gastric mucosa and particularly the gastrin cell. J Mol Endocrin 20: 111–117. 10.1677/jme.0.0200111
- Waldum HL, Sandvik AK, Idle JR (1998). Gastrin is the most important factor in ECL tumorigenesis. Gastroenterology 114: 1113–1115. 10.1016/s0016-5085(98)70346-4
- Williams DA (1971). A test for differences between treatment means when several dose levels are compared with a zero-dose control. Biometrics 27: 103–117. https://doi.org/10.2307/2528930
- Williams DA (1972). The comparison of several dose levels with a zero-dose control. Biometrics 28: 519–531. https://doi.org/10.2307/2556164
- Zhao CM, Chen D, Lintunen M, Paula P, Hakanson R (1999). Secretory organelles in ECL cells of the rat stomach: An immunohistochemical and electron-microscopic study. Cell Tissue Res 298(3): 457–470. 10.1007/s004419900075